New volumes of the Encyclopædia Britannica > Volume 30, K-MOR
(844) Page 798

Download files
Complete book:
Individual page:
{kind=link}
Thumbnail gallery: Grid view | List view

798
MOLLUSCA
rounded by four circles of cilia, to form the velum. The blasto¬
pore remains open, and draws gradually towards the anterior end
along the ventral surface. On the dorsal aspect spring two lateral
pallial ridges, parallel and symmetrical; these stretch towards the
ventral surface, where they finally fuse to form a tubular mantle
round the body. The shell which this mantle secretes, at first
cup-shaped, little by little assumes the form of a tube, like the
mantle, by union of its margins. On the ventral surface the foot
appears as a prominence ; this becomes elongated anteriorly, and
by its aid the animal can crawl after the disappearance of the
velum. The cerebral ganglia arise as two ectodermal invagina¬
tions in the velar area ; the otocysts by invagination at the surface
of the foot ; and the pedal ganglia, after the otocysts, by ecto¬
dermal thickenings. The anus is perforated very late. After five
or six days the velum atrophies, and the embryo begins to crawl.
Families: Dentaliidse, Siphonodentalihke.
Class IY. LamellibrancMa. — The old classification, founded
on the number of adductor muscles, can no longer be retained,
the “Monomya” being polyphyletic. The degree of specializa¬
tion attained by different forms is well shown by the structure
of the branchiae (ctenidia); using these, five orders may be
distinguished.
Order 1. Profo&nrocMz.—Branchial filaments short, not flexed,
not united by junctions, arranged in two series in opposite direc¬
tions. Families : Nuculidse, Solcnomyuke.
Order 2. —Branchial filaments elongated, parallel,
flexed, and united together by ciliary junctions. Families:
Anomiidse, Arcidse, Trigoniidse, Mytilidae.
Order 3. Pseudolamellibranchia.—Branchial filaments with
interfoliate junctions (vascular or connective); branchiae plicated ;
posterior adductor muscle reduced or wanting; foot slightly
developed. Families: Aviculidae, Ostrseidae, Pectinidse, Dimyidae.
Order 4. Eulamellibranchia. — Branchiae with interfilamentar
and interfoliar unions, all of which are vascular ; mantle lobes
united posteriorly at one or two points. This order includes the
great majority of families, which are distributed among seven sub¬
orders : Submytilacea, Tellinacea, Veneracea Cardiacea, Myacea,
Pholadacea, and Anatinacea.
Order 5. Septibranchia.—Branchiae transformed into a muscular
septum, which stretches from the anterior adductor to the
separation of the two siphons, and surrounds the foot. Families.:
Poromyidae, Cuspidariidae.
As in other Mollusca, the edges of the mantle can be reflected
over (Galeomma, Entovalva), or can even entirely conceal the shell,
which is absolutely internal in three genera, Chlamydoconcha,
Ephippodonta, and Scioberetia. In the last the adductor
muscles are absent, as in Aspergillum. Both adductors are
embryonically developed in all Lamellibranchs, the anterior ap-
p°arin~ rd.-.v.yc firct, even in cases where it ultimately disappears.
The redactor muscles of the foot correspond to the columellar
muscle of Gastropoda. The foot has a ventral disc only in
Protobranchia and Pectunculus ; in other forms it exhibits very
generally a cavity, the cells of which secrete a byssus for fixation
of the animal. The liver occasionally presents an asymmetry, the
left lobe being in such cases the larger (Nuculidse). The epithelial
covering of the auricles often constitutes pericardial glands ; other
portions of the wall of the pericardium may penetrate into the
mantle. In the Septibranchia, by abnormal development of their
muscular elements, the branchiae form a muscular septum with
symmetrical orifices ; respiration is effected by the internal surface
of the mantle, over which the contractions of the septum force a
powerful current of water. The two nephridia communicate with
one another; they extend sometimes into the mantle. The
genital glands also communicate with one another in many forms ;
and in some cases open into the kidneys. Many species are
hermaphrodite, but while in some (e.g., Ostraea, Cardium) ova
and spermatozoa arise side by side throughout the extent of
the gland, in others {e.g., Pecten) male and female portions are
■distinct ; and in Poromya and Anatinacea there exist a pair of
ovaries and a pair of testes entirely distinct, each with its proper
orifice.
In the more archaic forms (Protobranchia) the pleural ganglia
are distinct, as in Gastropoda and Scaphopoda; they are coupled
to the cerebral, each giving rise to a pallial nerve, and a pleuro-
pedal connective more or less promptly united to a cerebro-pedal
connective. The existence of the pleural ganglia shows that no
other visceral centre is fused with the cerebral, and that these are
the posterior centres which correspond to the visceral ganglia of
other Mollusca. The otocysts remain open to the exterior in some
adult Nuculidse. The cephalic eyes observable in various larvae
are retained in some adults on the anterior region of the branchial
axis (Mytilus, Avicula, Fig. 7). A unique structure characterizes
the pallial eyes of Pectinidar and of some species of Cardium.
The ova are laid separately (except in ISTucula delphinodonta).
A large number of Lamellibranchia retain the developing eggs in
the branchial lamellae (Submytilaoea, and fresh-water forms,
except Dreissensi). Larval nephridia have been detected in several
sub-groups, consisting of a deep canalicular portion, and a super¬
ficial portion which opens to the exterior postero-ventrally of the
head ; the lumen is intracellular, with a flame cell.
In the Unionidae the larva (Glochidium) is temporarily parasitic,
and encysts in the skin of a fish, deriving sustenance by means of
the ectodermal cells of the
mantle from the epidermis of
the host. Parasitic life lasts
from two to five weeks.
Class Y. Cephalopoda. — For
definitions of the divisions, the
article in vol. xvi. of this work
should be consulted.
Subclass A. Tetrabranchia.
—This contains a single living
genus, Nautilus, and numerous
fossil genera (see article Cepha¬
lopoda).
Subclass B. Dibranchia.
Order 1. Decapoda.—Suborder
{a) QDgopsida, with seven living
families: Spiral idee, Ommato-
strephidse, Physanoteuthidse,
Onychoteuthidse, Gonatidse,
Chiroteuthidse, Cranchiidse.
Suborder {b) Myopsida. Families
riidse, Loliginidse, Sepiidae
Order 2. Odopoda.—Families
Argonautidse, Philonexidse.
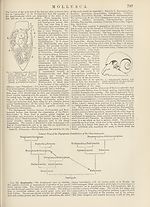
Fig. 7.—Avicula, left view of anterior
part of the body : pa, anterior palp;
ve, foot; pp, posterior palp; by,
byssus; br, internal gill-plate ; 6r'2,
external gill-plate ; m, reflected left
mantle lobe; o, eye (on the first
internal gill filament).
: Sepiolidse, Idiosepiidae, Sepiida-
Cirrhoteuthidse, Octopodidse,
The internal shell is greatly reduced ;
it is purely chitinous in all Octopoda, forming an unpaired piece
in Cirrhoteuthis, a pair of stylets in Octopus. In Argonauta alone
the shell gland shallows out, without closing. In the Decapoda some
allies of Sepiola are also shell-less. The tentacular arms atrophy
in various (Egopsida. The hectocotylized arm belongs generally
to the fourth pair in Decapoda, that of the left side only (Ommato-
strephidae, Onychoteuthidae, and most Myopsida), or on both
sides (Idiosepius, Spirula, Fig. 8). In Nautilus the spadix may be
right or left. Some deep-water (Egopsida possess luminous organs.
The radula is absent from Cirrhoteuthis ; the ink-sac is also want¬
ing in this genus and in some species of Octopus. In Octopoda
the pericardial part of the coelom is wanting. The coelom includes
only the genital capsule, which communicates with the capsules
of the branchial hearts by long canals (absent in two families).
The appendices of the branchial hearts are the morphological
equivalent of the “pericardial glands”
of other Mollusca ; their glandular wall
is excretory. The nervous system presents
the same labial commissure as do Amphi-
neura and Prorhipidoglossomorpha. The
osphradiurr. of ^utilus is placed against
and protected by the interbranchial
papilla. Certain pigmented organs of
Cirrhoteuthis are supposed to be thermo-
scopic. In those Octopoda in which the
hectocotyl us is not autotomous, it is
intruded into the pallial cavity of the
female so as to introduce spermatophores
into the distal portion of the oviduct
(Octopus) ; both arms of the dorsal pair
in Sepiolidae place the spermatophores
in the neighbourhood of the female open¬
ing.
In Nautilus the ova, which are four or
five millimetres long, are laid separately
in two shells, the outer of which is
partially opened. In the Dibranchia,
the nerve-centres arise from ectodermal
thickenings; the brachial ganglia are
formed by subdivision of the primitive
Fl(Huxley rad^Pelsemee?): Pedal g^Slia’ thus showing the pedal
F, F1, fins; Pd, dorsal origin o: the arms. 1 he external portion
anterior mantle-lobe; T, of the crystalline lens has a separate
right tentacular arm ; origin from the internal portion. The
I,’II, III, IF, right arms.’ blastoderm of the egg forms the ecto¬
derm ; the primitive endoderm takes
origin at the periphery of the ectoderm, spreading under and
away from it, enclosing the yolk with a layer of large nuclei at its
surface to form the perivitelline membrane. The remainder of the
endoderm is in great measure transformed into the mesoderm. The
primitive endoderm being unable to surround itself with ectoderm
to form a gastrula, the definitive endoderm appears rather late in
development; a furrow of cells below the hinder part of the mantle
in the middle line, derived, like the perivitelline membrane, from
the primitive endoderm, gives rise to stomach, liver, and intestine.
Authorities.—1. Von Jhering. Vergleichende Anatomic des
Nervensystemes und Phylogenie der Mollusken, Leipzig, 1877.—2.
Bouvier, “Systeme nerveux, morphologic genera'le et classifica*
MOLLUSCA
rounded by four circles of cilia, to form the velum. The blasto¬
pore remains open, and draws gradually towards the anterior end
along the ventral surface. On the dorsal aspect spring two lateral
pallial ridges, parallel and symmetrical; these stretch towards the
ventral surface, where they finally fuse to form a tubular mantle
round the body. The shell which this mantle secretes, at first
cup-shaped, little by little assumes the form of a tube, like the
mantle, by union of its margins. On the ventral surface the foot
appears as a prominence ; this becomes elongated anteriorly, and
by its aid the animal can crawl after the disappearance of the
velum. The cerebral ganglia arise as two ectodermal invagina¬
tions in the velar area ; the otocysts by invagination at the surface
of the foot ; and the pedal ganglia, after the otocysts, by ecto¬
dermal thickenings. The anus is perforated very late. After five
or six days the velum atrophies, and the embryo begins to crawl.
Families: Dentaliidse, Siphonodentalihke.
Class IY. LamellibrancMa. — The old classification, founded
on the number of adductor muscles, can no longer be retained,
the “Monomya” being polyphyletic. The degree of specializa¬
tion attained by different forms is well shown by the structure
of the branchiae (ctenidia); using these, five orders may be
distinguished.
Order 1. Profo&nrocMz.—Branchial filaments short, not flexed,
not united by junctions, arranged in two series in opposite direc¬
tions. Families : Nuculidse, Solcnomyuke.
Order 2. —Branchial filaments elongated, parallel,
flexed, and united together by ciliary junctions. Families:
Anomiidse, Arcidse, Trigoniidse, Mytilidae.
Order 3. Pseudolamellibranchia.—Branchial filaments with
interfoliate junctions (vascular or connective); branchiae plicated ;
posterior adductor muscle reduced or wanting; foot slightly
developed. Families: Aviculidae, Ostrseidae, Pectinidse, Dimyidae.
Order 4. Eulamellibranchia. — Branchiae with interfilamentar
and interfoliar unions, all of which are vascular ; mantle lobes
united posteriorly at one or two points. This order includes the
great majority of families, which are distributed among seven sub¬
orders : Submytilacea, Tellinacea, Veneracea Cardiacea, Myacea,
Pholadacea, and Anatinacea.
Order 5. Septibranchia.—Branchiae transformed into a muscular
septum, which stretches from the anterior adductor to the
separation of the two siphons, and surrounds the foot. Families.:
Poromyidae, Cuspidariidae.
As in other Mollusca, the edges of the mantle can be reflected
over (Galeomma, Entovalva), or can even entirely conceal the shell,
which is absolutely internal in three genera, Chlamydoconcha,
Ephippodonta, and Scioberetia. In the last the adductor
muscles are absent, as in Aspergillum. Both adductors are
embryonically developed in all Lamellibranchs, the anterior ap-
p°arin~ rd.-.v.yc firct, even in cases where it ultimately disappears.
The redactor muscles of the foot correspond to the columellar
muscle of Gastropoda. The foot has a ventral disc only in
Protobranchia and Pectunculus ; in other forms it exhibits very
generally a cavity, the cells of which secrete a byssus for fixation
of the animal. The liver occasionally presents an asymmetry, the
left lobe being in such cases the larger (Nuculidse). The epithelial
covering of the auricles often constitutes pericardial glands ; other
portions of the wall of the pericardium may penetrate into the
mantle. In the Septibranchia, by abnormal development of their
muscular elements, the branchiae form a muscular septum with
symmetrical orifices ; respiration is effected by the internal surface
of the mantle, over which the contractions of the septum force a
powerful current of water. The two nephridia communicate with
one another; they extend sometimes into the mantle. The
genital glands also communicate with one another in many forms ;
and in some cases open into the kidneys. Many species are
hermaphrodite, but while in some (e.g., Ostraea, Cardium) ova
and spermatozoa arise side by side throughout the extent of
the gland, in others {e.g., Pecten) male and female portions are
■distinct ; and in Poromya and Anatinacea there exist a pair of
ovaries and a pair of testes entirely distinct, each with its proper
orifice.
In the more archaic forms (Protobranchia) the pleural ganglia
are distinct, as in Gastropoda and Scaphopoda; they are coupled
to the cerebral, each giving rise to a pallial nerve, and a pleuro-
pedal connective more or less promptly united to a cerebro-pedal
connective. The existence of the pleural ganglia shows that no
other visceral centre is fused with the cerebral, and that these are
the posterior centres which correspond to the visceral ganglia of
other Mollusca. The otocysts remain open to the exterior in some
adult Nuculidse. The cephalic eyes observable in various larvae
are retained in some adults on the anterior region of the branchial
axis (Mytilus, Avicula, Fig. 7). A unique structure characterizes
the pallial eyes of Pectinidar and of some species of Cardium.
The ova are laid separately (except in ISTucula delphinodonta).
A large number of Lamellibranchia retain the developing eggs in
the branchial lamellae (Submytilaoea, and fresh-water forms,
except Dreissensi). Larval nephridia have been detected in several
sub-groups, consisting of a deep canalicular portion, and a super¬
ficial portion which opens to the exterior postero-ventrally of the
head ; the lumen is intracellular, with a flame cell.
In the Unionidae the larva (Glochidium) is temporarily parasitic,
and encysts in the skin of a fish, deriving sustenance by means of
the ectodermal cells of the
mantle from the epidermis of
the host. Parasitic life lasts
from two to five weeks.
Class Y. Cephalopoda. — For
definitions of the divisions, the
article in vol. xvi. of this work
should be consulted.
Subclass A. Tetrabranchia.
—This contains a single living
genus, Nautilus, and numerous
fossil genera (see article Cepha¬
lopoda).
Subclass B. Dibranchia.
Order 1. Decapoda.—Suborder
{a) QDgopsida, with seven living
families: Spiral idee, Ommato-
strephidse, Physanoteuthidse,
Onychoteuthidse, Gonatidse,
Chiroteuthidse, Cranchiidse.
Suborder {b) Myopsida. Families
riidse, Loliginidse, Sepiidae
Order 2. Odopoda.—Families
Argonautidse, Philonexidse.
Fig. 7.—Avicula, left view of anterior
part of the body : pa, anterior palp;
ve, foot; pp, posterior palp; by,
byssus; br, internal gill-plate ; 6r'2,
external gill-plate ; m, reflected left
mantle lobe; o, eye (on the first
internal gill filament).
: Sepiolidse, Idiosepiidae, Sepiida-
Cirrhoteuthidse, Octopodidse,
The internal shell is greatly reduced ;
it is purely chitinous in all Octopoda, forming an unpaired piece
in Cirrhoteuthis, a pair of stylets in Octopus. In Argonauta alone
the shell gland shallows out, without closing. In the Decapoda some
allies of Sepiola are also shell-less. The tentacular arms atrophy
in various (Egopsida. The hectocotylized arm belongs generally
to the fourth pair in Decapoda, that of the left side only (Ommato-
strephidae, Onychoteuthidae, and most Myopsida), or on both
sides (Idiosepius, Spirula, Fig. 8). In Nautilus the spadix may be
right or left. Some deep-water (Egopsida possess luminous organs.
The radula is absent from Cirrhoteuthis ; the ink-sac is also want¬
ing in this genus and in some species of Octopus. In Octopoda
the pericardial part of the coelom is wanting. The coelom includes
only the genital capsule, which communicates with the capsules
of the branchial hearts by long canals (absent in two families).
The appendices of the branchial hearts are the morphological
equivalent of the “pericardial glands”
of other Mollusca ; their glandular wall
is excretory. The nervous system presents
the same labial commissure as do Amphi-
neura and Prorhipidoglossomorpha. The
osphradiurr. of ^utilus is placed against
and protected by the interbranchial
papilla. Certain pigmented organs of
Cirrhoteuthis are supposed to be thermo-
scopic. In those Octopoda in which the
hectocotyl us is not autotomous, it is
intruded into the pallial cavity of the
female so as to introduce spermatophores
into the distal portion of the oviduct
(Octopus) ; both arms of the dorsal pair
in Sepiolidae place the spermatophores
in the neighbourhood of the female open¬
ing.
In Nautilus the ova, which are four or
five millimetres long, are laid separately
in two shells, the outer of which is
partially opened. In the Dibranchia,
the nerve-centres arise from ectodermal
thickenings; the brachial ganglia are
formed by subdivision of the primitive
Fl(Huxley rad^Pelsemee?): Pedal g^Slia’ thus showing the pedal
F, F1, fins; Pd, dorsal origin o: the arms. 1 he external portion
anterior mantle-lobe; T, of the crystalline lens has a separate
right tentacular arm ; origin from the internal portion. The
I,’II, III, IF, right arms.’ blastoderm of the egg forms the ecto¬
derm ; the primitive endoderm takes
origin at the periphery of the ectoderm, spreading under and
away from it, enclosing the yolk with a layer of large nuclei at its
surface to form the perivitelline membrane. The remainder of the
endoderm is in great measure transformed into the mesoderm. The
primitive endoderm being unable to surround itself with ectoderm
to form a gastrula, the definitive endoderm appears rather late in
development; a furrow of cells below the hinder part of the mantle
in the middle line, derived, like the perivitelline membrane, from
the primitive endoderm, gives rise to stomach, liver, and intestine.
Authorities.—1. Von Jhering. Vergleichende Anatomic des
Nervensystemes und Phylogenie der Mollusken, Leipzig, 1877.—2.
Bouvier, “Systeme nerveux, morphologic genera'le et classifica*
Set display mode to:
![]() Universal Viewer |
Universal Viewer | ![]() Mirador |
Large image | Transcription
Mirador |
Large image | Transcription
Images and transcriptions on this page, including medium image downloads, may be used under the Creative Commons Attribution 4.0 International Licence unless otherwise stated. ![]()
| Encyclopaedia Britannica > New volumes of the Encyclopædia Britannica > Volume 30, K-MOR > (844) Page 798 |
|---|
| Permanent URL | https://digital.nls.uk/193578473 |
|---|
| Attribution and copyright: |
|
|---|---|
| Shelfmark | EB.18 |
|---|---|
| Description | Ten editions of 'Encyclopaedia Britannica', issued from 1768-1903, in 231 volumes. Originally issued in 100 weekly parts (3 volumes) between 1768 and 1771 by publishers: Colin Macfarquhar and Andrew Bell (Edinburgh); editor: William Smellie: engraver: Andrew Bell. Expanded editions in the 19th century featured more volumes and contributions from leading experts in their fields. Managed and published in Edinburgh up to the 9th edition (25 volumes, from 1875-1889); the 10th edition (1902-1903) re-issued the 9th edition, with 11 supplementary volumes. |
|---|---|
| Additional NLS resources: |
|